最終氷期最寒冷期のトウヒ属球果について (吉川昌伸(2016) を一部抜粋・改変)
日本に自生するトウヒ属は、トウヒ節のエゾマツとトウヒ、バラモミ節のアカエゾマツ、ヒメバラモミ(=アズサバラモミ)、ヤツガタケトウヒ(=ヒメマツハダ)、ハリモミ、イラモミの7種である(佐竹他編1989;Katsuki et.al.2004)。トウヒ節の分布は、エゾマツは北海道および北東アジア、トウヒはエゾマツの変種であり本州中部の亜高山帯である(佐竹他編1989)。バラモミ節の各種の分布は、アカエゾマツは北海道に分布の中心がありサハリン最南端や岩手県早池峰山、イラモミは福島県南部から岐阜県までの山地帯上部から亜高山帯、ヒメバラモミとヤツガタケトウヒは本州中部の八ヶ岳と南アルプス北部等などの山地帯上部から亜高山帯、ハリモミは他種と異なり山地帯に生え福島県以南の太平洋側で多く日本海側の多雪地には殆ど分布しない(佐竹1989;(独法)森林総合研究所森林遺伝研究領域2011)。トウヒ属の球果は、トウヒ節とバラモミ節で区別でき、バラモミ節のうちハリモミは分布が異なるため、他のバラモミ節4種で識別できるかどうかが問題になる。
最終氷期最寒冷期に、関東地方や東北地方に分布していたトウヒ属の種構成は明らかでない。鈴木・竹内(1989)は仙台市富沢遺跡から出土したトウヒ属の球果を当初はアカエゾマツとした。しかし、その後、富沢遺跡から産出した約1,000個の球果の形態変異から、アカエゾマツに近似するが現生種と鱗片の特徴が異なるためヤツガタケトウヒに近似な絶滅種のトミザワトウヒとし、一部球果はアズサバラモミに近似な絶滅種のコウシントウヒとした(鈴木1992)。小西・鈴木(1997)は、現生アカエゾマツとヤツガタケトウヒの球果形態の変異に基づき、トミザワトウヒはアカエゾマツの変異に含まれるとし、種鱗の先端の波打ちが無いことによりアカエゾマツから区別して別種とすることは出来ないこと、球果化石のサイズに依存した同定を見直す必要があることを示した。また、野手ほか(1998)は論文等に記載された球果や種鱗のサイズと種鱗先端の形態に基づき、トウヒ属バラモミ節樹木を球果形態から複数種に区分することは困難であるとした。小西・鈴木(1997)と野手ほか(1998)は球果や種鱗のサイズと種燐先端の波打ちの程度に基づき識別できないことを示したが、種鱗の厚さの変化や種鱗表面の模様は評価されていないため、これら研究による球果形態の検討は十分とは言えない。
Kobayashi et al.(2000)は葉緑体DNAの塩基配列に基づき、津軽半島出来島海岸のAT下位の泥炭層から出土した球果をアカエゾマツ、新郷村間明田の八戸テフラ直下から採取した針葉をヤツガタケトウヒとし、さらに球果形態による同定とDNAでの区別が一致しており、トウヒ属の一部は球果形態で種を識別できる可能性を示した。トミザワトウヒは、種鱗表面に縦方向のしわが目立ち、種鱗の縁がやや薄くなる特徴があり(吉川1995)、現生種ではヤツガタケトウヒに似るとされる(鈴木1992)。現生のアカエゾマツは種鱗先端の波打ちの程度が全く平滑なものから激しく波打つものまである(小西・鈴木1997)。出来島海岸から産出したアカエゾマツには種鱗に波打ちが認められるが(Kobayashi et al.,2000)、トミザワトウヒの種鱗には波打ちが認められる個体が産出していないことから(鈴木1992;吉川1995)、トミザワトウヒはアカエゾマツとは別種である可能性がある。
一方で、ヤツガタケトウヒ近似種とされた球果が、東京都中野区北江古田遺跡の立川期(ATより上位で約22,400 14C yr BP)と、栃木県南部の二宮町の立川期(約20,290
14 C yr BP)から産出している。江古田遺跡の球果は、球果サイズと種鱗の形からアカエゾマツの球果変異の中に含まれるとする見解があるが(小西・鈴木1997)、前述のように球果形態の検討が十分でないため支持できない。東北地方北部からヤツガタケトウヒが産出し、富沢遺跡のトミザワトウヒがヤツガタケトウヒに似ること、関東ではヤツガタケトウヒに似た球果が産出している状況からは、ヤツガタケトウヒが関東から東北地方北部までの低地に連続的に分布していた可能性を示唆している。現状ではアカエゾマツは青森県の日本海側で確認されたものの、ヤツガタケトウヒとアカエゾマツの分布は明らかでない。今後、球果形態の再検討と球果のDNAも並行して検討する必要がある。
最終氷期最盛期には、東北地方南部まで北方系針葉樹が南下したことが強調されてきた。しかし、現在、中部地方に分布するやチョウセンゴヨウが関東地方から東北地方の低地の広い範囲に分布し(津村・百原2011)、東北地方北部からヤツガタケトウヒが産出しており、関東地方から東北地方の低地の広範囲に本州中部系針葉樹が分布していた可能性がある。
ところで、人の植物利用からみるとチョウセンゴヨウの種子は可食できるものの、アカエゾマツやヤツガタケトウヒの利用価値はほとんどなかったかもしれません。
引用文献
独立行政法人森林総合研究所森林遺伝研究領域.2011.希少樹種の現状と保全―保全のための課 題と対策―.https://www.ffpri.affrc.go.jp/labs/raretree/3_PKMindex.html、2015年11月引用
Katsuki,T., Takeuchi,T., Sugaya,K., Kitamuta,T., Katsuta,M., Yoshimura,H.
2004. Geographic distribution and genetic variation of a vulnerable conifer
species, Picea koyamae (Pinaceae). Acta Phytotax .Geebot..55 :19-28
Kobayashi, K., Yoshikawa,J., Suzuki, M. 2000. DNA identification of Picea
species of the Last Glacial Age in northem Japan.. Jpn. J. Histor. Botany
2:67-80
小西彰一・鈴木三男.1997.アカエゾマツの球果形態の変異.植生史研究5:67-76
能城修一・鈴木三男.1992.仙台市富沢遺跡から出土した木材化石の樹種と森林植生の復元.「富沢遺跡‐第30次調査報告書第Ⅱ分冊‐」:231-243頁、仙台市、仙台市教育委員会
野手啓行・沖津進・百原新.1998.日本のトウヒ属バラモミ節樹木の現在の分布と最終氷期以後の分布変遷.植生史研究6:3-13
佐竹義輔・原 寛・亘理俊次・冨成忠夫.1989. 日本の野生植物 木本Ⅰ.321頁、東京、平凡社
鈴木敬治.1992.大型植物化石.「富沢遺跡‐第30次調査報告書第Ⅱ分冊‐」:244-273、仙台市、仙台市教育委員会
鈴木敬治・竹内貞子.1989.中~後期更新世における古植物相―東北地方を中心として―.第四紀研究28:303-316
津村義彦・百原 新.2011.植物化石とDNAからみた温帯性針樹木の最終氷期最盛期のレフュージア.「日本列島の三万五千年-人と自然の環境史 第6巻 環境史をとらえる技法」(湯本貴和編):59-75頁、東京、文一総合出版
吉川純子.1995.仙台市富沢遺跡第88次調査で産出した大型植物化石.「富沢遺跡第88次・89次発掘調査報告書第」:50-67頁、仙台市、仙台市教育委員会
(吉川昌伸.2016.更新世末から完新世初頭の東北日本の植生史. 旧石器研究第12号:1-12)
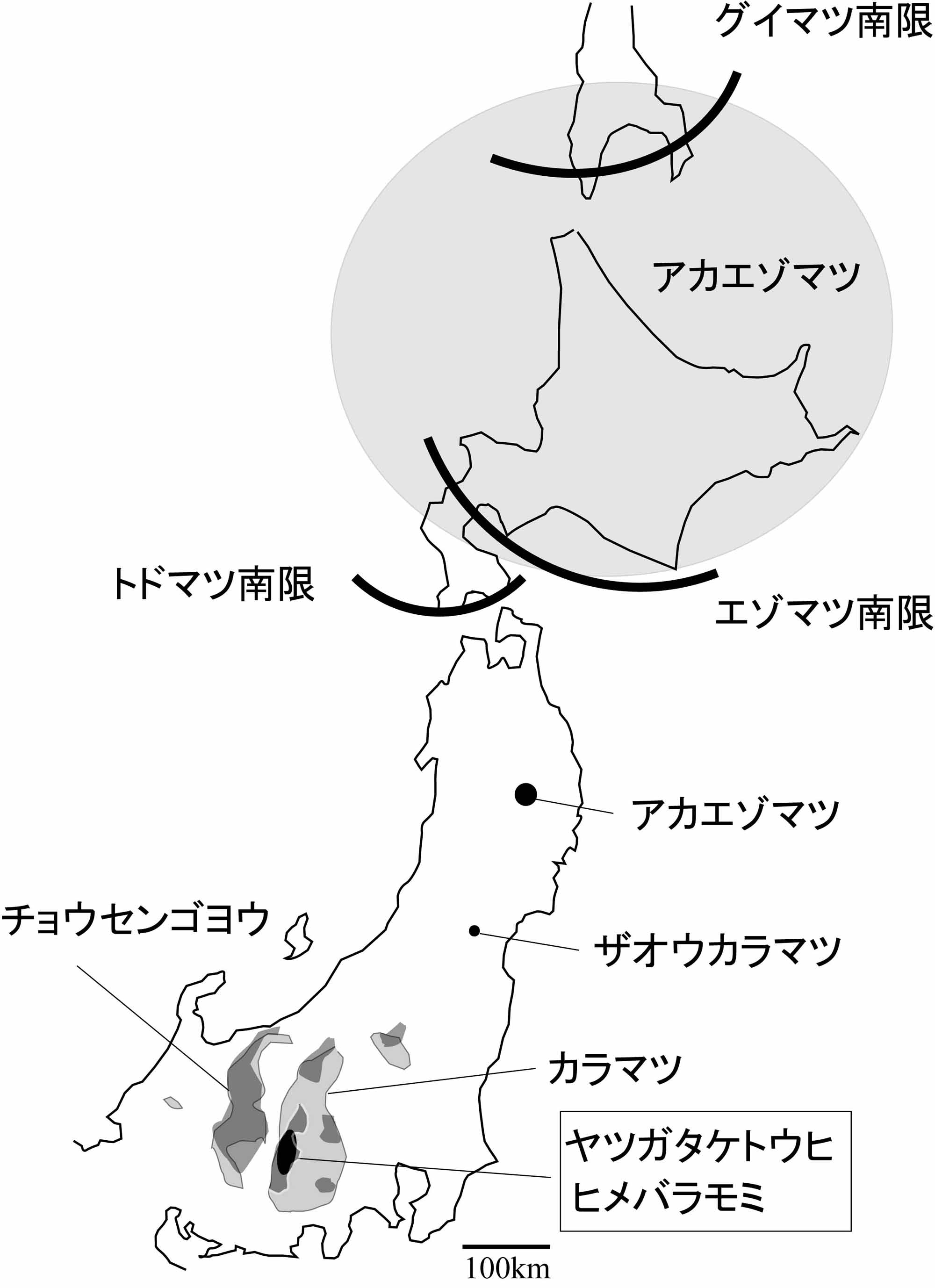 亜高山と亜寒帯針葉樹の現在の分布
亜高山と亜寒帯針葉樹の現在の分布
アカエゾマツ ヒメバラモミ ヤツガタケトウヒ